Wahoooo it’s online!!! 🎉🥳
Excited to share a glimpse into my postdoc work with @chorye.bsky.social at Duke where @stefan-golas.bsky.social and I developed TurboPRANCE, a robotics platform to scale and enable new phage-assisted continuous evolutions. bskytorial party!👇 1/n
04.03.2026 15:24
👍 10
🔁 3
💬 1
📌 0
Redirecting
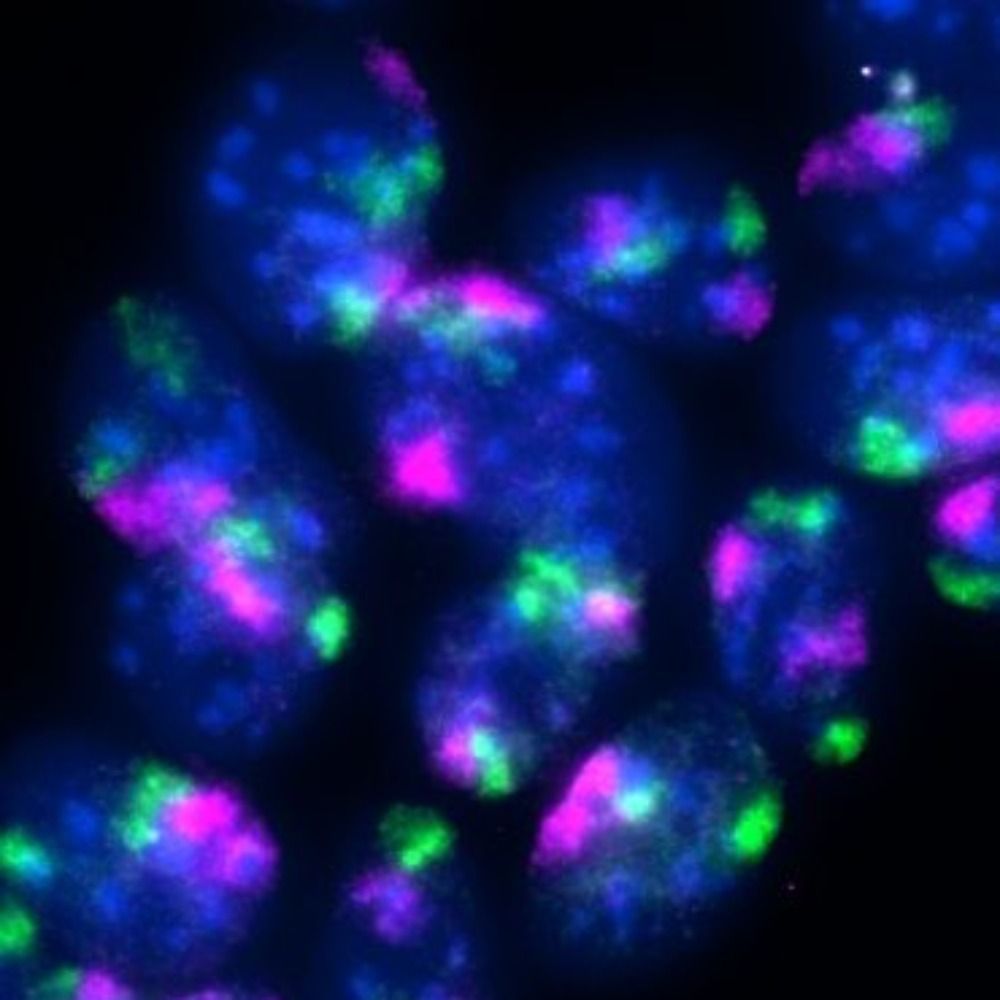
Our most recent work on the “function and evolution” of #nuclear-speckles is now online at Cell @cp-cell.bsky.social
doi.org/10.1016/j.ce...
Read the thread👇 for the highlights of our findings.
25.02.2026 16:01
👍 122
🔁 59
💬 9
📌 5

Chromatin spatial analysis by METALoci unveils sex-determining 3D regulatory hubs
Nature Structural & Molecular Biology - METALoci, a new three-dimensional genome computational tool, reveals a major rewiring of regulatory interactions during sex determination. By combining...
🤩💪 Out now! 3D regulatory hubs in sex determination
With @mamartirenom.bsky.social & Capel labs, led by @imotagom.bsky.social & @jrotwitguez.bsky.social
1️⃣ METALoci – explore #3DGenome 🧬
2️⃣ Non-coding region controlling Fgf9 🧩
3️⃣ Meis genes = new key players 🌟
👉 rdcu.be/e5sm2
1/n Bluetorial 👇
24.02.2026 14:02
👍 57
🔁 26
💬 1
📌 2


Excited to share @suminkim.bsky.social and @mileshuseyin.bsky.social 's new Current Opinion review on how Polycomb complexes mediate 3D genome interactions including mechanistic models and potential roles in gene regulation:
www.sciencedirect.com/science/arti...
18.02.2026 22:03
👍 60
🔁 18
💬 1
📌 2
24/ This work was led by multi-talented @erikacanderson.bsky.social
with expert contributions from:
Hadi Rahmaninejad, @gfudenberg.bsky.social
@aljahani.bsky.social , Ivana Cavka, Alistair Boettiger
@emilyarnold.bsky.social , Annie Adachi,
@rinishah.bsky.social
@karissalhansen.bsky.social
12.02.2026 21:56
👍 3
🔁 0
💬 0
📌 0
23/ Altogether genomic context shapes the local flux of loop extrusion within which enhancers operate.
So the next big question now becomes how cells control general mechanisms of looping to tune long-range enhancer action… 🧐
12.02.2026 21:56
👍 2
🔁 0
💬 1
📌 0

22/ Interestingly, Enformer, a transformer-based machine learning model, predicts that the same CTCF site retains less cohesin when it is flanked by other CTCF sites.
> Current ML models learned rules for CTCF-CTCF interference from genomic context, despite being unaware of loop extrusion.
12.02.2026 21:56
👍 3
🔁 0
💬 1
📌 0

21/ Combining simulations and ChIP-seq, we found that a CTCF site interferes with how much cohesin its neighbor can receive.
This interference patern explains variation in cohesin levels at CTCF sites genome-wide, without the need to invoke targeted loading at privileged elements like enhancers.
12.02.2026 21:56
👍 0
🔁 0
💬 1
📌 0

20/ The cohesin blocked at CTCF sites gives clues about where extrusion happens -- some CTCF sites retain way more cohesin that others, meaning the cohesin flux is not uniform across the genome.
12.02.2026 21:56
👍 1
🔁 0
💬 1
📌 0

19/ So the key question now becomes: what controls the flux of cohesin extrusion in the first place, if not targeted loading at enhancers?
Through careful ChIP-seq background subtraction we found cohesin is largely (~75%) between CTCF sites.
12.02.2026 21:56
👍 1
🔁 0
💬 1
📌 0

18/ All in all, we do not find evidence that enhancers load cohesin to support their own long-range activity on distal promoter targets.
Instead, enhancer action appears subordinate to the general flux of cohesin extrusion.
12.02.2026 21:56
👍 5
🔁 0
💬 1
📌 0

17/ 🖥️> Enhancers generally do not display such ‘jets’/’fountains’, at least in normal mouse ES cells. Yet cohesin can help them act long-range. How??
We find alternative regimes where enhancers may still modulate cohesin flux but without acting as loading sites, by tuning loading across their TAD
12.02.2026 21:56
👍 1
🔁 0
💬 1
📌 0

16/ 🖥️> Weaker targeted loading creates an antidiagonal on Hi-C maps (=‘jets’/’fountains’), but this does not increase interaction between the loading site and the rest of the domain.
i.e. targeted loading at enhancers does not by itself increase their chance to interact with distal promoters.
12.02.2026 21:56
👍 0
🔁 0
💬 1
📌 0
15/ 🖥️> In simulations, very strong targeted loading over several kilobases creates boundaries and stripes on contact maps. Yet enhancers generally do not display such patterns in Hi-C/micro-C.
12.02.2026 21:48
👍 0
🔁 0
💬 1
📌 0

14/🖥️> loading at enhancers would need to be rather unrealistic to overcome the flux of cohesin coming from the background and enrich cohesin even just 2x at nearby CTCF sites
(i.e. if your enhancer KO disrupts cohesin binding, it is unlikely to be just because of targeted loading at the enhancer)
12.02.2026 21:48
👍 0
🔁 0
💬 1
📌 0

13/ Many calculations later 📐📏:
🖥️> cohesin is not expected to be retained at loading sites
(i.e. if you see cohesin ChIP-seq at enhancers, it not likely to be a signature of targeted loading)
12.02.2026 21:48
👍 1
🔁 0
💬 1
📌 0

12/ Hadi Rahmaninejad in the group of @gfudenberg.bsky.social did just that, using biophysical simulations of loop extrusion. 🖥️
12.02.2026 21:48
👍 1
🔁 0
💬 1
📌 0

11/ 🤔…
Let’s reconsider our expectations of targeted cohesin loading then.
Enhancers represent such a small fraction of te genome, they would need to boost cohesin loading REALLY REALLY high to outweigh background loading rate.
Can we put numbers on this to help interpret experimental results?
12.02.2026 21:48
👍 0
🔁 0
💬 1
📌 0

10/ 🔬Chromosome tracing microscopy by @aljahani.bsky.social & Ivana Cavka in Alistair Boettiger’s group showed that targeted cohesin loading disrupts local 3D folding, creating a boundary.
Yet the negative impact on enhancer function must involve additional effects (cohesin::RNA-pol/TF clashes?).
12.02.2026 21:48
👍 2
🔁 0
💬 1
📌 0
9/ Nope. ❌
Quite the opposite in fact.
Using the TArgeted Cohesin Loader (TACL) approach (www.nature.com/articles/s41...) from Wouter de Laat’s group to increase loading was very detrimental to distal transcription.
So more cohesin loading by the enhancer =/= better enhancer function
12.02.2026 21:43
👍 2
🔁 0
💬 1
📌 0
8/ Let’s attack the problem more directly, from the functional angle.
Is increasing cohesin loading by an enhancer beneficial to its long-range activity? 📝
12.02.2026 21:42
👍 0
🔁 0
💬 1
📌 0


7/ Using an artificial tethering system we saw that transcription activators are sufficient to recruit NIPBL/MAU2. Yet we don’t see cohesin being recruited…
Does that mean NIPBL/MAU2 at enhancer does not load cohesin then? 😵💫
12.02.2026 21:42
👍 0
🔁 0
💬 1
📌 0


6/ Using new quantitative controls with clean tags, inducible degrons and calibrated ChIP-seq, we saw that NIPBL/MAU2 (cohesin cofactor complex necessary for initiation and progression of loop extrusion) binds at enhancers in mouse ES cells. Yet we don’t see much cohesin (RAD21)…
12.02.2026 21:41
👍 0
🔁 0
💬 1
📌 0
5/ Targeted vs. uniform loading models have VERY different implications for how we think about transcriptional regulation and enhancer functions.
So, what is happening in cells?
12.02.2026 21:41
👍 0
🔁 0
💬 1
📌 0

4/ Contrasting with this view is the idea that cohesin loads uniformly, and that enhancers are instead subordinate to the general dynamics of cohesin looping. 🤔
12.02.2026 21:39
👍 1
🔁 0
💬 1
📌 0

3/ This would mean that enhancers have some autonomy in setting up the DNA loops that support their role in long-range transcriptional regulation 💪
12.02.2026 21:39
👍 0
🔁 0
💬 1
📌 0
2/ Enhancers have been proposed to act as privileged loading sites for cohesin, raising the interesting idea that they actively fold the genome to engage distal target promoters and activate them.
12.02.2026 21:39
👍 0
🔁 0
💬 1
📌 0
Interested in transcriptional regulation, enhancers and 3D genome folding?
In this new study we wondered about the role of cohesin loading at enhancers for long-range transcriptional control
www.biorxiv.org/content/10.6...
detailed 🧵👇
12.02.2026 21:39
👍 67
🔁 33
💬 1
📌 3
Bay Area Chromatin Club - In Person Symposium: March 5, 2026
The Chromatin Club Bay Area was founded to foster a scientific community between academic and industry members in and around the Bay Area.
We are starting Bay Area Chromatin Club up again in 2026! Fist event March 5th at UCSF Mission Bay! Details and registration HERE: If you like to nominate a person for a talk- DM me. It's been a great experience in the past.
urldefense.com/v3/__https:/...
09.02.2026 21:40
👍 9
🔁 7
💬 0
📌 1